Frontiers in Human Neuroscience (Dansk)
Introduktion
Bevidsthed er en ophidselse og bevidsthed om miljø og selv, som opnås gennem handling af det stigende retikulære aktiveringssystem (ARAS) på hjernestammen og hjernebarken (Daube, 1986; Paus, 2000; Zeman, 2001; Gosseries et al., 2011). ARAS består af flere neuronale kredsløb, der forbinder hjernestammen til cortex. Disse neuronale forbindelser stammer hovedsageligt fra hjernestammens retikulære dannelse (RF) og projicerer gennem synaptiske relæer i den intralaminære kerne af thalamus til hjernebarken (Daube, 1986; Paus, 2000; Zeman, 2001; Afifi og Bergman, 2005; Gosseries et al., 2011). Derudover er adskillige hjernestammekerner (locus coeruleus, dorsal raphe, median raphe, pedunculopontine nucleus, parabrachial nucleus), ikke-specifikke thalamiske kerner, hypothalamus og basal forhjerne også inkluderet i ARAS-systemet (Aston-Jones et al., 2001 ; Parvizi og Damasio, 2003; Fuller et al., 2011). Grundig evaluering af ARAS er vigtig for diagnose og behandling af patienter med nedsat bevidsthed, såsom patienter, der er i en vegetativ tilstand eller dem med minimal bevidsthed (Zeman, 2001; Gosseries et al., 2011).
Konventionel MR-hjerne, funktionelle neuroimaging-teknikker, elektrofysiologiske metoder og MR-spektroskopi er blevet anvendt i studier af ARAS i den menneskelige hjerne (Parvizi og Damasio, 2003; Schiff, 2006; Tshibanda et al., 2009, 2010; Gawryluk et al. ., 2010). Men fordi ARAS ikke klart kan diskrimineres fra tilstødende neurale strukturer, kan nøjagtig identifikation og estimering af ARAS i den menneskelige hjerne være problematisk, når man bruger disse metoder. I modsætning hertil tillader diffusion tensor imaging (DTI) evaluering af hvidt stof på grund af dets evne til at afbilde vanddiffusionsegenskaber (Mori et al., 1999). I normalt hvidt stof har vandmolekyler relativ bevægelsesfrihed parallelt med nervefiberskanaler. Imidlertid er deres bevægelser begrænset på tværs af kanaler, hvilket giver anledning til diffusionsanisotropi af hvidt stof. Derfor er diffusionsanisotropi blevet anvendt til evaluering af omfanget af fiberændring i hvidt stof (Chang et al., 2010; Puig et al., 2010). Flere nylige undersøgelser har forsøgt at demonstrere nytten af DTI til evaluering af læsioner hos patienter med nedsat bevidsthed og tilslutning af specifikke ARAS-kerner i den menneskelige hjerne (Voss et al., 2006; Perlbarg et al., 2009; Tollard et al., 2009; Tshibanda et al., 2009; Fernandez-Espejo et al., 2010, 2011; Newcombe et al., 2010; Edlow et al., 2012). Imidlertid vides der ikke meget om hele rekonstruktionen og estimeringen af ARAS i den menneskelige hjerne (Edlow et al., 2012).
I den aktuelle undersøgelse, ved hjælp af DTI, forsøgte vi at rekonstruere den nedre enkeltkomponent. af ARAS fra pontin RF til de intralaminære kerner i thalamus i den normale menneskelige hjerne.
Materialer og metoder
Emner
26 normale raske forsøgspersoner (14 mænd, 12 kvinder; gennemsnitsalder, 31,85 ± 9,80 år; interval, 20–50) uden historie med neurologisk sygdom blev rekrutteret til denne undersøgelse. Alle forsøgspersoner deltog i denne undersøgelse som frivillige og gav skriftligt samtykke, inden de gennemgik DTI-scanning. Undersøgelsen blev godkendt af det institutionelle revisionsudvalg på vores hospital.
Diffusion Tensor Image
DTI-data blev erhvervet ved hjælp af en 6-kanals hovedspole på en 1,5-T Philips Gyroscan Intera ( Philips, Best, Holland) med single-shot ekko-plan billeddannelse. For hver af de 32 ikke-kollinære diffusionssensibiliserende gradienter erhvervede vi 67 sammenhængende skiver parallelt med den forreste commissure-posterior commissure line. Billeddannelsesparametre var som følger: erhvervelsesmatrix = 96 × 96, rekonstrueret til matrix = 128 × 128, synsfelt = 221 mm × 221 mm, TR = 10.726 ms, TE = 76 ms, parallel billedreduktionsfaktor (SENSE-faktor) = 2, EPI-faktor = 49, og b = 1000 s / mm2, NEX = 1, og en skivetykkelse på 2,3 mm (erhvervet isotrop voxelstørrelse 2,3 mm × 2,3 mm × 2,3 mm).
Probabilistisk fiber Sporing
Analyse af diffusionsvægtede billeddata blev udført ved hjælp af Oxford Center for Functional Magnetic Resonance Imaging of the Brain (FMRIB) softwarebibliotek (FSL; www.fmrib.ox.ac.uk/fsl). Affine multi-skala to-dimensionel registrering blev brugt til korrektion af hovedbevægelseseffekt og billedforvrængning på grund af virvelstrøm. Fibersporing blev udført ved hjælp af en probabilistisk traktografimetode baseret på en multifibermodel og anvendt i den aktuelle undersøgelse under anvendelse af traktografirutiner implementeret i FMRIB Diffusion (5000 strømlineprøver, trinlængder på 0,5 mm, krumningstærskler = 0,2) (Smith et al., 2004 ).Fordele ved probabilistisk traktografi, som blev brugt i denne undersøgelse, inkluderer større robusthed over for støj såvel som evnen til at detektere stier med skarpere vinkler og at skelne krydsende fibre (Behrens et al., 2007; Winston et al., 2011).
ARAS-stien blev bestemt ved udvælgelse af fibre, der passerer gennem frøområder af interesse (ROI) og mål (termination) ROIer. Et frø-ROI blev placeret på RF af ponserne på niveauet for trigeminusnervenindgangszonen (Daube, 1986; Afifi og Bergman, 2005). Analyse af den mediale lemniscus og rubrospinal kanal blev udført for at bekræfte grænsen for RF på pons (figur 1A). Til analyse af den mediale lemniscus blev frø-ROIer placeret på den anteromediale medulla, og mål-ROI blev placeret på den somatosensoriske cortex (Hong et al., 2010). Til analyse af rubrospinalkanalen blev frø-ROIer placeret på den røde kerne, og mål-ROI blev placeret på den kontralaterale dorsolaterale region i medulla (Monakow-området) (Nathan og Smith, 1982; Kwon et al., 2011). Målet ROI blev givet på thalamusens intralaminære kerner på niveauet for kommissærplanet (Morel, 2007). Ved definitionen af thalamusens intralaminære kerner henviste vi til et hjerneatlas (Morel, 2007) (figur 1A). Af 5000 prøver genereret fra frøvoxelen blev resultater for kontakt visualiseret ved en tærskel på mindst 1 strømlinet gennem hver voxel til analyse. Værdier for fraktioneret anisotropi (FA), gennemsnitlig diffusivitet (MD) og kanalnummer for den nedre enkeltkomponent af ARAS blev målt.
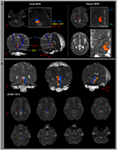
Figur 1. (A) Frøområder af interesse (ROI) er angivet på den pontine retikulære formation (rød farve). Mål-ROI er angivet på thalamusens intralaminære kerner på niveauet for kommissærplanet. Grænsen for thalamusens intralaminære kerner blev defineret ved henvisning til lærebogen i hjerneatlaset (Morel, 2007). ML, medial lemniscus; RST, rubrospinal kanal; RF, retikulær dannelse; AC, forreste kommission PC, bageste kommission. (B) Veje for det rekonstruerede stigende retikulære aktiveringssystem er vist på hvert niveau af hjernen i et normalt subjekt (26-årig mand).
Statistisk analyse
SPSS-software (v.15.0; SPSS, Chicago, IL, USA) blev anvendt til dataanalyse. Parret t-test blev anvendt til bestemmelse af forskellen i værdier af ARIs DTI-parametre mellem højre og venstre halvkugle. Pearson-korrelationstest blev anvendt til bestemmelse af korrelation mellem DTI-parametre for ARAS og alder. Resultater blev betragtet som signifikante, når p-værdien var < 0,05.
Resultater
Vi rekonstruerede den nedre enkeltkomponent i ARAS mellem pontinen RF og intralaminar kerner i thalamus. Den rekonstruerede komponent af ARAS stammer fra pontin RF, steg op gennem det mesencephalic tegmentum lige bagud til den røde kerne og afsluttedes derefter på de intralaminære kerner i thalamus på niveauet for kommissurplanet i alle forsøgspersoner (figur 1B). / p>
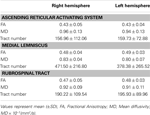
Tabel 1. Diffusionstensor-billeddannelsesparametre i det stigende retikulære aktiveringssystem .
Diskussion
I den aktuelle undersøgelse, ved hjælp af DTI, rekonstruerede vi en af hovedveje for ARAS, den nedre enkeltkomponent af ARAS fra RF til thalamus hos normale forsøgspersoner, skønt ARAS består af yderligere hjernestammekerner, hypothalamus, basal forhjerne og thalamocorticale fremspring til hjernebarken. Vi valgte to ROIer til rekonstruktion af den nederste enkeltkomponent i ARAS: frø-ROI, som var RF for pons på niveauet for trigeminusnervenindgangszonen (Daube, 1986; Afifi og Bergman, 2005) og målet ROI, som omfattede thalamusens intralaminære kerner (de centrale laterale kerner, centromedian / parafascikulære kerner og paracentrale kerner) på niveauet med kommissærplanet (Morel, 2007). Den rostrale del af RF i hjernestammen over trigeminusnerven indgangszone er kendt som ARAS; derimod er den kaudale del af RF involveret i motorfunktion og autonom funktion relateret til hjerte- og åndedrætsfunktion (Daube, 1986). Derfor placerede vi frø-ROI i RF på niveauet for trigeminusnervens indgangszone. Vi placerede mål-ROI i de intralaminære kerner, som er de vigtigste kerner i ARAS blandt de ikke-specifikke thalamiske kerner. Derfor mener vi, at fordi vi ikke kunne inkludere de andre thalamiske kerner, der er involveret i ARAS, er den nedre enkeltkomponent i ARAS, der blev rekonstrueret i den aktuelle undersøgelse, ikke hele den nedre enkeltkomponent i ARAS, men hoveddelen af hele den nedre enkeltkomponent i ARAS.Derfor stammede den nedre enkeltkomponent i ARAS fra pontin RF, steg op gennem det mesencephalic tegmentum bageste til den røde kerne og sluttede derefter på thalamusens intralaminære kerner. Derudover adskilt værdierne for FA, MD og kanalnumre for den rekonstruerede nedre enkeltkomponent i ARAS ikke signifikant mellem højre og venstre halvkugle. Kanalnummeret bestemmes af antallet af voxels indeholdt i en neural kanal (Kwak et al., 2010). FA-værdien indikerer graden af retningsbestemmelse og integritet af hvide substansmikrostrukturer såsom axoner, myelin og mikrotubuli, og ADC-værdien angiver størrelsen af vanddiffusion (Assaf og Pasternak, 2008).
Flere undersøgelser har demonstreret den kliniske anvendelighed af DTI ved at estimere nogle områder af den nedre enkeltkomponent i ARAS fra RF til thalamus hos patienter med nedsat bevidsthed (Perlbarg et al., 2009; Tollard et al., 2009; Newcombe et al., 2010; Fernandez-Espejo et al., 2011). Tollard et al. (2009) rapporterede om nytten af DTI, som blev udført på det subakutte stadium til forudsigelse af resultatet hos 45 patienter med svær TBI (traumatisk hjerneskade) (fravær af respons på enkle ordrer). I deres undersøgelse målte de FA-værdien i flere supratentorielle og infratentoriale områder, inklusive de forreste pons, posterior pons og midthjernen og viste, at faldet i infratentorial og supratentorial FA, undtagen i posterior pons, tillod forudsigelse af ugunstige resultater 1 år fra TBI. Perlbarg et al. (2009), der udførte DTI-scanning hos 30 patienter med fravær af respons på enkle ordrer efter svær TBI, rapporterede et bestemt fald i FA målt i den underordnede langsgående fasciculus, midthjernen (cerebral peduncle og tegmentum), bageste lem af den indre kapsel og bageste corpus callosum i den ugunstige resultatgruppe. Newcombe et al. (2010) brugte DTI til karakterisering af omfanget og placeringen af tab af hvidt stof hos patienter, der var i en vegetativ tilstand sekundært til TBI (syv patienter) og patienter med iskæmisk-hypoxisk skade (fem patienter). Abnormaliteter i de supratentorale områder blev observeret i begge grupper; derimod blev abnormiteter i hjernestammen kun observeret i TBI-gruppen. Fernandez-Espejo et al. (2011) brugte DTI til differentiering af neuropatologien hos 25 vegetative og minimalt bevidste patienter. De konkluderede, at patienter med minimal bevidsthed og dem, der var i vegetativ tilstand, var forskellige i subkortisk hvid substans og thalamiske regioner, men syntes ikke at være forskellige i hjernestammen. I en nylig undersøgelse under anvendelse af diffusionsbilleddannelse med høj vinkelopløsning, Edlow et al. (2012) rapporterede om neuroanatomisk forbindelse af ARAS i den menneskelige hjerne, både in vivo og ex vivo. De demonstrerede, at forbindelserne mellem specifikke ARAS-kerner var impliceret i ophidselse, og de af thalamiske kerner var impliceret i modulering af ophidselse.
Afslutningsvis rekonstruerede vi den nedre enkeltkomponent i ARAS fra DTI RF til thalamus i den menneskelige hjerne. Vi mener, at den anvendte metode og resultaterne af denne undersøgelse kan være nyttige for forskere, der studerer ARAS i den menneskelige hjerne. En af begrænsningerne ved denne undersøgelse er imidlertid, at vi ikke var i stand til fuldt ud at belyse hele ARAS-systemet, fordi vi ikke inkluderede andre thalamiske og hjernestamme kerner i vores analyse, som også er involveret i ARAS. Yderligere undersøgelser af den kliniske anvendelighed af vores fund samt undersøgelser af fremskrivningerne af ARAS fra thalamus til hjernebarken er nødvendige.
Erklæring om interessekonflikt
Forfatterne erklærer at forskningen blev udført i fravær af kommercielle eller økonomiske forhold, der kunne opfattes som en potentiel interessekonflikt.
Anerkendelser
Dette arbejde blev støttet af DGIST R & D Program fra Koreas ministerium for uddannelse, videnskab og teknologi (13-BD-0401).
Afifi, AK og Bergman, RA (2005). Funktionel neuroanatomi: tekst og atlas. New York: Lange Medical Books / McGraw-Hill.
Morel, A. (2007). Stereotaktisk atlas over menneskelig thalamus og basalganglier. New York: Informa Healthcare.