Frontiers in Human Neuroscience (Norsk)
Introduksjon
Bevissthet er en opphisselse og bevissthet om miljø og selvtillit, som oppnås gjennom handling av det stigende retikulære aktiveringssystemet (ARAS) på hjernestammen og hjernebarken (Daube, 1986; Paus, 2000; Zeman, 2001; Gosseries et al., 2011). ARAS består av flere nevronale kretser som forbinder hjernestammen med hjernebarken. Disse nevronale forbindelsene har hovedsakelig sin opprinnelse i retikulær formasjon (RF) i hjernestammen og projiserer gjennom synaptiske reléer i den intralaminære kjernen i thalamus til hjernebarken (Daube, 1986; Paus, 2000; Zeman, 2001; Afifi og Bergman, 2005; Gosseries et al., 2011). I tillegg er flere hjernestammekjerner (locus coeruleus, dorsal raphe, median raphe, pedunculopontine nucleus, parabrachial nucleus), ikke-spesifikke thalamiske kjerner, hypothalamus og basal forhjernen inkludert i ARAS-systemet (Aston-Jones et al., 2001 ; Parvizi og Damasio, 2003; Fuller et al., 2011). Grundig evaluering av ARAS er viktig for diagnose og behandling av pasienter med nedsatt bevissthet, for eksempel pasienter som er i vegetativ tilstand eller de med minimal bevissthet (Zeman, 2001; Gosseries et al., 2011).
Konvensjonell hjerne-MR, funksjonelle nevroavbildningsteknikker, elektrofysiologiske metoder og MR-spektroskopi har blitt brukt i studier av ARAS i den menneskelige hjerne (Parvizi og Damasio, 2003; Schiff, 2006; Tshibanda et al., 2009, 2010; Gawryluk et al. ., 2010). Men fordi ARAS ikke kan diskrimineres tydelig fra tilstøtende nevrale strukturer, kan nøyaktig identifisering og estimering av ARAS i den menneskelige hjerne være problematisk når du bruker disse metodene. I kontrast tillater diffusjonstensoravbildning (DTI) evaluering av hvitt materiale på grunn av dets evne til å avbilde vanndiffusjonsegenskaper (Mori et al., 1999). I normal hvit substans har vannmolekyler relativ bevegelsesfrihet parallelt med nervefiberkanaler. Imidlertid er deres bevegelser begrenset over traktater, noe som gir opphav til diffusjonsanisotropi av hvit materie. Følgelig har diffusjonsanisotropi blitt brukt for å evaluere omfanget av fiberendring i hvitt materiale (Chang et al., 2010; Puig et al., 2010). Flere nylige studier har forsøkt å demonstrere nytten av DTI for evaluering av lesjoner hos pasienter med nedsatt bevissthet og tilkobling av spesifikke ARAS-kjerner i den menneskelige hjerne (Voss et al., 2006; Perlbarg et al., 2009; Tollard et al., 2009; Tshibanda et al., 2009; Fernandez-Espejo et al., 2010, 2011; Newcombe et al., 2010; Edlow et al., 2012). Imidlertid er det lite kjent om hele rekonstruksjonen og estimeringen av ARAS i den menneskelige hjerne (Edlow et al., 2012).
I den nåværende studien, med DTI, forsøkte vi å rekonstruere den nedre enkeltkomponenten. av ARAS fra pontin RF til de intralaminære kjernene i thalamus i den normale menneskelige hjerne.
Materialer og metoder
Fag
Tjuefem normale friske forsøkspersoner (14 menn, 12 kvinner; gjennomsnittsalder, 31,85 ± 9,80 år; rekkevidde, 20–50) uten historie med nevrologisk sykdom ble rekruttert til denne studien. Alle fag deltok i denne studien som frivillige og ga skriftlig samtykke før de gjennomgikk DTI-skanning. Studien ble godkjent av det institusjonelle vurderingsnemnda på sykehuset vårt.
Diffusjonstensorbilde
DTI-data ble anskaffet ved hjelp av en 6-kanals hodespole på en 1,5-T Philips Gyroscan Intera ( Philips, Best, Nederland) med ekko-plan bildebehandling. For hver av de 32 ikke-kollinære diffusjonssensibiliserende gradientene, anskaffet vi 67 sammenhengende skiver parallelt med den fremre commissure-posterior commissure-linjen. Bildeparametere var som følger: oppkjøpsmatrise = 96 × 96, rekonstruert til matrise = 128 × 128, synsfelt = 221 mm × 221 mm, TR = 10 726 ms, TE = 76 ms, parallell reduksjonsfaktor for bildebehandling (SENSE-faktor) = 2, EPI-faktor = 49 og b = 1000 s / mm2, NEX = 1, og en skivetykkelse på 2,3 mm (ervervet isotrop voxelstørrelse 2,3 mm × 2,3 mm × 2,3 mm).
Probabilistic Fiber Sporing
Analyse av diffusjonsveide bildedata ble utført ved bruk av Oxford Center for Functional Magnetic Resonance Imaging of the Brain (FMRIB) Software Library (FSL; www.fmrib.ox.ac.uk/fsl). Affine flerskala todimensjonal registrering ble brukt for å korrigere hodebevegelseseffekt og bildeforvrengning på grunn av virvelstrøm. Fibersporing ble utført ved bruk av en sannsynlig traktografimetode basert på en multifibermodell, og anvendt i den nåværende studien ved bruk av traktografirutiner implementert i FMRIB Diffusion (5000 strømlinjeformede prøver, trinnlengder på 0,5 mm, krumningsterskler = 0,2) (Smith et al., 2004 ).Fordeler med sannsynlig traktografi, som ble brukt i denne studien, inkluderer større robusthet mot støy, samt evnen til å oppdage stier med skarpere vinkler og å skille kryssende fibre (Behrens et al., 2007; Winston et al., 2011).
ARAS-banen ble bestemt ved valg av fibre som passerer gjennom frøområder av interesse (ROI) og mål (terminering) ROI. En ROI av frø ble plassert på RF av ponsene på nivået av trigeminusnervens inngangssone (Daube, 1986; Afifi og Bergman, 2005). Analyse av medial lemniscus og rubrospinal kanal ble utført for å bekrefte grensen for RF på ponsene (figur 1A). For analyse av medial lemniscus ble frø-ROIer plassert på anteromedial medulla og mål-ROI ble plassert på den somatosensoriske cortexen (Hong et al., 2010). For analyse av rubrospinalkanalen ble frø-ROI plassert på den røde kjernen, og mål-ROI ble plassert på den kontralaterale dorsolaterale regionen i medulla (Monakow’s area) (Nathan og Smith, 1982; Kwon et al., 2011). Målet ROI ble gitt på de intralaminare kjernene i thalamus på nivået med kommisjonsplanet (Morel, 2007). Ved å definere thalamusens intralaminære kjerner, refererte vi til et hjerneatlas (Morel, 2007) (figur 1A). Av 5000 prøver generert fra seed voxel ble resultatene for kontakt visualisert med en terskel på minimum 1 strømlinjeformet gjennom hver voxel for analyse. Verdier av fraksjonell anisotropi (FA), gjennomsnittlig diffusivitet (MD) og kanalnummer for den nedre enkeltkomponenten av ARAS ble målt.
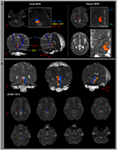
Figur 1. (A) Frøområder av interesse (ROI) er gitt på den pontine retikulære formasjonen (rød farge). Målavkastningen er gitt på de intralaminære kjernene i thalamus på nivået med kommisjonsplanet. Grense for thalamusens intralaminære kjerner ble definert med henvisning til læreboken til hjerneatlaset (Morel, 2007). ML, medial lemniscus; RST, rubrospinal kanal; RF, retikulær formasjon; AC, fremre kommisjon PC, bakre kommisjon. (B) Veier til det rekonstruerte stigende retikulære aktiveringssystemet er vist på hvert nivå av hjernen i et normalt subjekt (26 år gammel mann).
Statistisk analyse
SPSS-programvare (v.15.0; SPSS, Chicago, IL, USA) ble brukt til dataanalyse. Paret t-test ble brukt for å bestemme forskjellen i verdier av DTI-parametere for ARAS mellom høyre og venstre halvkule. Pearson korrelasjonstest ble brukt for å bestemme korrelasjon mellom DTI-parametere for ARAS og alder. Resultatene ble ansett som signifikante når p-verdien var < 0,05.
Resultater
Vi rekonstruerte den nedre enkeltkomponenten i ARAS mellom pontinen RF og intralaminar kjerner i thalamus. Den rekonstruerte komponenten av ARAS stammer fra pontine RF, steg opp gjennom mesencephalic tegmentum like bakre til den røde kjernen, og avsluttet deretter på de intralaminære kjernene i thalamus på nivået med kommisjonsplanet i alle fag (Figur 1B). / p>
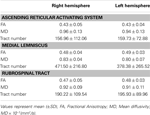
Tabell 1. Diffusjonstensoravbildningsparametere til det stigende retikulære aktiveringssystemet .
Diskusjon
I den nåværende studien, ved hjelp av DTI, rekonstruerte vi en av hovedveiene til ARAS, den nedre enkeltkomponenten av ARAS fra RF til thalamus hos normale individer, selv om ARAS består av ekstra hjernestammekjerner, hypothalamus, basal forhjerne og thalamocortical projeksjoner til hjernebarken. Vi valgte to ROIer for rekonstruksjon av den nedre enkeltkomponenten i ARAS: frø-ROI, som var RF for ponsene på nivået av trigeminusnervens inngangssone (Daube, 1986; Afifi og Bergman, 2005), og målet ROI, som inkluderte thalamusens intralaminære kjerner (de sentrale laterale kjerner, centromedian / parafascikulære kjerner og paracentrale kjerner) på nivået med kommisjonsplanet (Morel, 2007). Den rostrale delen av RF av hjernestammen over trigeminusnerven inngangssone er kjent som ARAS; derimot er den kaudale delen av RF involvert i motorisk funksjon og autonom funksjon relatert til hjerte- og respirasjonsfunksjon (Daube, 1986). Derfor plasserte vi frø-ROI i RF på nivået av trigeminusnervens inngangssone. Vi plasserte målavkastningen i de intralaminare kjernene, som er hovedkjernene til ARAS blant de ikke-spesifikke thalamykjernene. Derfor tror vi at fordi vi ikke kunne inkludere de andre thalamykjernene som er opptatt av ARAS, er den nedre enkeltkomponenten av ARAS som ble rekonstruert i den nåværende studien ikke hele den nedre enkeltkomponenten i ARAS, men hoveddelen av hele den nedre enkeltkomponenten i ARAS.Følgelig stammer den nedre enkeltkomponenten av ARAS fra pontin RF, steg opp gjennom mesencephalic tegmentum bakre til den røde kjernen, og ble deretter avsluttet på de intralaminære kjernene i thalamus. I tillegg skilte ikke verdiene for FA, MD og traktatnumrene til den rekonstruerte nedre enkeltkomponenten i ARAS seg signifikant mellom høyre og venstre halvkule. Kanalnummeret bestemmes av antall vokser som finnes i en nevral kanal (Kwak et al., 2010). FA-verdien indikerer graden av retningsretning og integritet av hvite substansmikrostrukturer som aksoner, myelin og mikrotubuli, og ADC-verdien indikerer størrelsen på vanndiffusjon (Assaf og Pasternak, 2008).
Flere studier har demonstrert den kliniske nytten av DTI ved å estimere noen områder av den nedre enkeltkomponenten i ARAS fra RF til thalamus hos pasienter med nedsatt bevissthet (Perlbarg et al., 2009; Tollard et al., 2009; Newcombe et al., 2010; Fernandez-Espejo et al., 2011). Tollard et al. (2009) rapporterte om nytten av DTI, som ble utført på det subakutte stadiet for å forutsi resultatet hos 45 pasienter med alvorlig TBI (traumatisk hjerneskade) (fravær av respons på enkle ordrer). I studien målte de FA-verdien i flere supratentorielle og infratentoriale områder, inkludert de fremre ponsene, de bakre ponsene og midthjernen, og demonstrerte at reduksjonen av infratentorial og supratentorial FA, bortsett fra i posterior pons, tillot forutsigelse av ugunstige resultater 1 år fra TBI. Perlbarg et al. (2009), som utførte DTI-skanning hos 30 pasienter med fravær av respons på enkle ordrer etter alvorlig TBI, rapporterte en klar reduksjon i FA målt i den underordnede langsgående fasciculus, mellomhjernen (cerebral peduncle og tegmentum), bakre lem av den indre kapselen , og posterior corpus callosum i den ugunstige resultatgruppen. Newcombe et al. (2010) brukte DTI for karakterisering av omfanget og plasseringen av tap av hvitt stoff hos pasienter som var i vegetativ tilstand sekundært til TBI (syv pasienter) og pasienter med iskemisk-hypoksisk skade (fem pasienter). Avvik i de supratentorielle områdene ble observert i begge gruppene; derimot ble abnormiteter i hjernestammen bare observert i TBI-gruppen. Fernandez-Espejo et al. (2011) brukte DTI i differensiering av nevropatologien til 25 vegetative og minimalt bevisste pasienter. De konkluderte med at pasienter med minimalt bevissthet og de som var i vegetativ tilstand, var forskjellige i subkortikale hvite substanser og thalamiske regioner, men syntes ikke å være forskjellige i hjernestammen. I en nylig studie med diffusjonsavbildning med høy vinkeloppløsning, Edlow et al. (2012) rapporterte om neuroanatomisk tilkobling av ARAS i den menneskelige hjerne, både in vivo og ex vivo. De demonstrerte at forbindelsene til spesifikke ARAS-kjerner var involvert i opphisselse, og de av thalamiske kjerner var implisert i modulering av opphisselse. RF til thalamus i den menneskelige hjerne. Vi tror at metoden som brukes og resultatene av denne studien kan være nyttig for forskere som studerer ARAS i den menneskelige hjerne. En av begrensningene i denne studien er imidlertid at vi ikke klarte å fullstendig belyse hele ARAS-systemet fordi vi ikke inkluderte andre thalamiske og hjernestamme kjerner i analysen vår som også er involvert i ARAS. Ytterligere studier av den kliniske nytten av våre funn, samt studier av fremskrivningene av ARAS fra thalamus til hjernebarken er nødvendig.
Erklæring om interessekonflikt
Forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kunne tolkes som en potensiell interessekonflikt.
Anerkjennelser
Dette arbeidet ble støttet av DGIST R & D-program fra Koreas utdannings-, vitenskaps- og teknologidepartement (13-BD-0401).
Afifi, AK og Bergman, RA (2005). Funksjonell neuroanatomi: tekst og atlas. New York: Lange Medical Books / McGraw-Hill.
Morel, A. (2007). Stereotaktisk atlas av menneskelig thalamus og basalganglia. New York: Informa Healthcare.