Frontières de la neuroscience humaine
Introduction
La conscience est une excitation et une prise de conscience de lenvironnement et de soi, qui est obtenue grâce à laction du système dactivation réticulaire ascendant (ARAS) sur le tronc cérébral et le cortex cérébral (Daube, 1986; Paus, 2000; Zeman, 2001; Gosseries et al., 2011). LARAS est composé de plusieurs circuits neuronaux reliant le tronc cérébral au cortex. Ces connexions neuronales proviennent principalement de la formation réticulaire (RF) du tronc cérébral et se projettent à travers des relais synaptiques dans le noyau intralaminaire du thalamus vers le cortex cérébral (Daube, 1986; Paus, 2000; Zeman, 2001; Afifi et Bergman, 2005; Gosseries et al., 2011). De plus, plusieurs noyaux du tronc cérébral (locus coeruleus, raphé dorsal, raphé médian, noyau pédonculopontin, noyau parabrachial), noyaux thalamiques non spécifiques, hypothalamus et cerveau antérieur basal sont également inclus dans le système ARAS (Aston-Jones et al., 2001 ; Parvizi et Damasio, 2003; Fuller et al., 2011). Une évaluation approfondie de lARAS est importante pour le diagnostic et la prise en charge des patients ayant une altération de la conscience, tels que les patients en état végétatif ou ceux ayant une conscience minimale (Zeman, 2001; Gosseries et al., 2011).
LIRM cérébrale conventionnelle, les techniques de neuroimagerie fonctionnelle, les méthodes électrophysiologiques et la spectroscopie RM ont été utilisées dans les études de lARAS dans le cerveau humain (Parvizi et Damasio, 2003; Schiff, 2006; Tshibanda et al., 2009, 2010; Gawryluk et al. ., 2010). Cependant, comme lARAS ne peut pas être clairement distingué des structures neuronales adjacentes, lidentification et lestimation précises de lARAS dans le cerveau humain peuvent être problématiques lors de lutilisation de ces méthodes. En revanche, limagerie par tenseur de diffusion (DTI) permet dévaluer la matière blanche en raison de sa capacité à imager les caractéristiques de diffusion de leau (Mori et al., 1999). Dans la matière blanche normale, les molécules deau ont une liberté de mouvement relative parallèle aux faisceaux de fibres nerveuses. Cependant, leurs mouvements sont limités à travers les étendues, ce qui donne lieu à une anisotropie de diffusion de la matière blanche. En conséquence, lanisotropie de diffusion a été utilisée pour évaluer lampleur du changement des fibres dans la substance blanche (Chang et al., 2010; Puig et al., 2010). Plusieurs études récentes ont tenté de démontrer lutilité du DTI pour lévaluation des lésions chez les patients ayant une conscience altérée et la connectivité de noyaux ARAS spécifiques dans le cerveau humain (Voss et al., 2006; Perlbarg et al., 2009; Tollard et al., 2009; Tshibanda et al., 2009; Fernandez-Espejo et al., 2010, 2011; Newcombe et al., 2010; Edlow et al., 2012). Cependant, on sait peu de choses sur lensemble de la reconstruction et de lestimation de lARAS dans le cerveau humain (Edlow et al., 2012).
Dans létude actuelle, en utilisant le DTI, nous avons tenté de reconstruire la composante unique inférieure de lARAS de la RF pontine aux noyaux intralaminaires du thalamus dans le cerveau humain normal.
Matériel et méthodes
Sujets
Vingt-six sujets sains normaux (14 hommes, 12 femmes; âge moyen, 31,85 ± 9,80 ans; intervalle de 20 à 50 ans) sans antécédent de maladie neurologique ont été recrutés pour cette étude. Tous les sujets ont participé à cette étude en tant que volontaires et ont fourni un consentement écrit avant de subir une analyse DTI. Létude a été approuvée par le comité dexamen institutionnel de notre hôpital.
Image du tenseur de diffusion
Les données DTI ont été acquises à laide dune bobine de tête à 6 canaux sur un Philips Gyroscan Intera de 1,5 T ( Philips, Best, Pays-Bas) avec imagerie écho-planaire à un seul coup. Pour chacun des 32 gradients de sensibilisation de diffusion non colinéaires, nous avons acquis 67 coupes contiguës parallèles à la ligne commissure antérieure-commissure postérieure. Les paramètres dimagerie étaient les suivants: matrice dacquisition = 96 × 96, reconstruite en matrice = 128 × 128, champ de vision = 221 mm × 221 mm, TR = 10726 ms, TE = 76 ms, facteur de réduction dimagerie parallèle (facteur SENSE) = 2, facteur EPI = 49 et b = 1000 s / mm2, NEX = 1, et une épaisseur de tranche de 2,3 mm (taille de voxel isotrope acquis 2,3 mm × 2,3 mm × 2,3 mm).
Fibre probabiliste Suivi
Lanalyse des données dimagerie pondérées en diffusion a été réalisée à laide de la bibliothèque logicielle du Centre dOxford pour limagerie par résonance magnétique fonctionnelle du cerveau (FMRIB) (FSL; www.fmrib.ox.ac.uk/fsl). Un enregistrement bidimensionnel multi-échelle affine a été utilisé pour la correction de leffet de mouvement de la tête et de la distorsion de limage due aux courants de Foucault. Le suivi des fibres a été réalisé à laide dune méthode de tractographie probabiliste basée sur un modèle multifibres, et appliqué dans létude actuelle en utilisant des routines de tractographie implémentées dans FMRIB Diffusion (5000 échantillons simplifiés, longueurs de pas de 0,5 mm, seuils de courbure = 0,2) (Smith et al., 2004 ).Les avantages de la tractographie probabiliste, qui a été utilisée dans cette étude, comprennent une plus grande robustesse au bruit, ainsi que la capacité de détecter des voies avec des angles plus nets et de distinguer les fibres qui se croisent (Behrens et al., 2007; Winston et al., 2011).
La voie de lARAS a été déterminée par la sélection de fibres passant par des régions de semences dintérêt (ROI) et des ROI cibles (de terminaison). Un ROI semencier a été placé sur le RF des pons au niveau de la zone dentrée du nerf trijumeau (Daube, 1986; Afifi et Bergman, 2005). Lanalyse du lemnisque médial et du tractus rubrospinal a été réalisée afin de confirmer la limite du RF sur les pontons (figure 1A). Pour lanalyse du lemnisque médial, des ROI de semences ont été placées sur la moelle antéro-médiale et la ROI cible a été placée sur le cortex somatosensoriel (Hong et al., 2010). Pour lanalyse du tractus rubrospinal, les ROI des semences ont été placées sur le noyau rouge et la ROI cible a été placée sur la région controlatérale dorsolatérale de la moelle (région de Monakow) (Nathan et Smith, 1982; Kwon et al., 2011). Le ROI cible a été donné sur les noyaux intralaminaires du thalamus au niveau du plan commissural (Morel, 2007). En définissant les noyaux intralaminaires du thalamus, nous nous sommes référés à un atlas du cerveau (Morel, 2007) (figure 1A). Sur 5000 échantillons générés à partir du voxel de graine, les résultats pour le contact ont été visualisés à un seuil minimum de 1 rationalisé à travers chaque voxel pour analyse. Les valeurs de lanisotropie fractionnaire (FA), de la diffusivité moyenne (MD) et du nombre de voies du composant unique inférieur dARAS ont été mesurées.
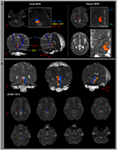
Figure 1. (A) Les régions de semences dintérêt (ROI) sont données sur la formation réticulaire pontine (couleur rouge). Le ROI cible est donné sur les noyaux intralaminaires du thalamus au niveau du plan commissural. La limite des noyaux intralaminaires du thalamus a été définie par référence au manuel de latlas du cerveau (Morel, 2007). ML, lemnisque médial; RST, tractus rubrospinal; RF, formation réticulaire; AC, commissure antérieure; PC, commissure postérieure. (B) Les voies du système dactivation réticulaire ascendant reconstruit sont représentées à chaque niveau du cerveau chez un sujet normal (homme de 26 ans).
Analyse statistique
Le logiciel SPSS (v.15.0; SPSS, Chicago, IL, USA) a été utilisé pour lanalyse des données. Un test t apparié a été utilisé pour déterminer la différence des valeurs des paramètres DTI de lARAS entre les hémisphères droit et gauche. Le test de corrélation de Pearson a été utilisé pour déterminer la corrélation entre les paramètres DTI de lARAS et lâge. Les résultats étaient considérés comme significatifs lorsque la valeur p était < 0,05.
Résultats
Nous avons reconstruit la composante simple inférieure de lARAS entre la pontine RF et noyaux intralaminaires du thalamus. Le composant reconstruit de lARAS provient du RF pontin, est monté à travers le tegmentum mésencéphalique juste en arrière du noyau rouge, puis sest terminé sur les noyaux intralaminaires du thalamus au niveau du plan commissural chez tous les sujets (Figure 1B).
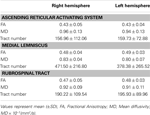
Tableau 1. Paramètres dimagerie du tenseur de diffusion du système dactivation réticulaire ascendant .
Discussion
Dans létude actuelle, en utilisant DTI, nous avons reconstruit lune des principales voies de lARAS, le composant unique inférieur de lARAS du RF au thalamus chez les sujets normaux, bien que lARAS se compose de noyaux supplémentaires du tronc cérébral, de lhypothalamus, du cerveau antérieur basal et des projections thalamocorticales vers le cortex cérébral. Nous avons sélectionné deux ROI pour la reconstruction de la composante unique inférieure de lARAS: le ROI de la graine, qui était le RF des pons au niveau de la zone dentrée du nerf trijumeau (Daube, 1986; Afifi et Bergman, 2005), et la cible ROI, qui incluait les noyaux intralaminaires du thalamus (noyaux latéraux centraux, noyaux centromédiens / parafasciculaires et noyaux paracentraux) au niveau du plan commissural (Morel, 2007). La partie rostrale du RF du tronc cérébral au-dessus de la zone dentrée du nerf trijumeau est connue sous le nom dARAS; en revanche, la partie caudale du RF est impliquée dans la fonction motrice et la fonction autonome liée à la fonction cardiaque et respiratoire (Daube, 1986). Par conséquent, nous avons placé le ROI de départ dans le RF au niveau de la zone dentrée du nerf trijumeau. Nous avons placé le ROI cible dans les noyaux intralaminaires, qui sont les noyaux principaux de lARAS parmi les noyaux thalamiques non spécifiques. Par conséquent, nous pensons que parce que nous ne pouvions pas inclure les autres noyaux thalamiques concernés par lARAS, le composant unique inférieur de lARAS qui a été reconstruit dans létude actuelle nest pas lensemble du composant unique inférieur de lARAS, mais la partie principale du tout composant unique inférieur de lARAS.Par conséquent, le composant unique inférieur de lARAS provenait du RF pontin, remontait à travers le tegmentum mésencéphalique postérieur au noyau rouge, puis se terminait sur les noyaux intralaminaires du thalamus. De plus, les valeurs des numéros FA, MD et des voies du composant unique inférieur reconstruit de lARAS ne différaient pas significativement entre les hémisphères droit et gauche. Le nombre de tractus est déterminé par le nombre de voxels contenus dans un tractus neural (Kwak et al., 2010). La valeur FA indique le degré de directionnalité et dintégrité des microstructures de matière blanche telles que les axones, la myéline et les microtubules, et la valeur ADC indique lampleur de la diffusion de leau (Assaf et Pasternak, 2008).
Plusieurs études ont démontré lutilité clinique du DTI en estimant certaines zones de la composante unique inférieure de lARAS, du RF au thalamus, chez des patients souffrant de troubles de la conscience (Perlbarg et al., 2009; Tollard et al., 2009; Newcombe et al., 2010; Fernandez-Espejo et al., 2011). Tollard et coll. (2009) ont rapporté lutilité du DTI, qui a été réalisé au stade subaigu pour la prédiction du résultat chez 45 patients atteints de TBI sévère (traumatisme crânien) (absence de réponse aux ordres simples). Dans leur étude, ils ont mesuré la valeur de FA au niveau de plusieurs zones supratentorielles et infratentorielles, y compris les pons antérieurs, postérieurs et mésencéphale, et ont démontré que la diminution de lAF infratentorielle et supratentorielle, sauf dans le pont postérieur, permettait de prédire des résultats défavorables. 1 an à partir de TBI. Perlbarg et coll. (2009), qui ont réalisé un scan DTI chez 30 patients en absence de réponse aux ordres simples suite à un TBI sévère, ont rapporté une nette diminution de lAF mesurée dans le fascicule longitudinal inférieur, le mésencéphale (pédoncule cérébral et tegmentum), le membre postérieur de la capsule interne et le corps calleux postérieur dans le groupe de résultat défavorable. Newcombe et coll. (2010) ont utilisé le DTI pour caractériser létendue et la localisation de la perte de substance blanche chez des patients qui étaient dans un état végétatif secondaire à un TBI (sept patients) et des patients présentant une lésion ischémique-hypoxique (cinq patients). Des anomalies dans les zones supratentorielles ont été observées dans les deux groupes; en revanche, des anomalies du tronc cérébral nont été observées que dans le groupe TBI. Fernandez-Espejo et coll. (2011) ont utilisé le DTI dans la différenciation de la neuropathologie de 25 patients végétatifs et minimalement conscients. Ils ont conclu que les patients peu conscients et ceux en état végétatif différaient dans la substance blanche sous-corticale et les régions thalamiques, mais ne semblaient pas différer dans le tronc cérébral. Dans une étude récente utilisant limagerie par diffusion à haute résolution angulaire, Edlow et al. (2012) ont décrit la connectivité neuroanatomique de lARAS dans le cerveau humain, à la fois in vivo et ex vivo. Ils ont démontré que les connectivités de noyaux ARAS spécifiques étaient impliquées dans lexcitation, et celles des noyaux thalamiques étaient impliquées dans la modulation de lexcitation.
En conclusion, en utilisant le DTI, nous avons reconstruit le composant unique inférieur de lARAS à partir du RF au thalamus dans le cerveau humain. Nous pensons que la méthodologie utilisée et les résultats de cette étude peuvent être utiles aux chercheurs qui étudient lARAS dans le cerveau humain. Cependant, lune des limites de cette étude est que nous navons pas pu élucider complètement lensemble du système ARAS car nous navons pas inclus dautres noyaux thalamiques et du tronc cérébral dans notre analyse qui sont également impliqués dans lARAS. Des études complémentaires sur lutilité clinique de nos résultats ainsi que des études sur les projections de lARAS du thalamus au cortex cérébral sont nécessaires.
Déclaration de conflit dintérêts
Les auteurs déclarent que la recherche a été menée en labsence de toute relation commerciale ou financière pouvant être interprétée comme un conflit dintérêts potentiel.
Remerciements
Ce travail a été soutenu par la DGIST R & Programme D du Ministère de léducation, de la science et de la technologie de Corée (13-BD-0401).
Afifi, AK et Bergman, RA (2005). Neuroanatomie fonctionnelle: texte et atlas. New York: Lange Medical Books / McGraw-Hill.
Morel, A. (2007). Atlas stéréotaxique du thalamus humain et des noyaux gris centraux. New York: Informa Healthcare.