Rajat ihmisen neurotieteessä
Johdanto
Tietoisuus on ympäristön ja itsensä kiihottuminen ja tietoisuus, joka saavutetaan nousevan verkkokalvon aktivoivan järjestelmän (ARAS) toiminnalla aivorungon ja aivokuoren (Daube, 1986; Paus, 2000; Zeman, 2001; Gosseries et ai., 2011). ARAS koostuu useista hermosolupiireistä, jotka yhdistävät aivorungon aivokuoreen. Nämä hermosoluyhteydet ovat peräisin pääasiassa aivorungon retikulaarisesta muodostumisesta (RF) ja heijastuvat talamuksen intralaminaarisessa ytimessä olevien synaptisten releiden kautta aivokuoreen (Daube, 1986; Paus, 2000; Zeman, 2001; Afifi ja Bergman, 2005; Gosseries et ai., 2011). Lisäksi ARAS-järjestelmään sisältyy myös useita aivorungon ytimiä (locus coeruleus, selkä raphe, mediaani raphe, pedunculopontine-ydin, parabrachial-ydin), epäspesifisiä talamuksen ytimiä, hypotalamusta ja tyvi-aivoja (Aston-Jones et al., 2001 ; Parvizi ja Damasio, 2003; Fuller ym., 2011). ARAS: n perusteellinen arviointi on tärkeää tajunnan vajaatoimintaa sairastavien potilaiden, kuten vegetatiivisessa tilassa olevien tai vähätajuisten potilaiden, diagnosoinnissa ja hoidossa (Zeman, 2001; Gosseries et al., 2011).
Perinteistä aivojen magneettikuvaa, toiminnallisia neurokuvantamistekniikoita, elektrofysiologisia menetelmiä ja MR-spektroskopiaa on käytetty ihmisen aivojen ARAS-tutkimuksissa (Parvizi ja Damasio, 2003; Schiff, 2006; Tshibanda et ai., 2009, 2010; Gawryluk et al. ., 2010). Koska ARAS: ta ei voida kuitenkaan selvästi erottaa vierekkäisistä hermorakenteista, ARAS: n tarkka tunnistaminen ja arviointi ihmisen aivoissa voi olla ongelmallista näitä menetelmiä käytettäessä. Sitä vastoin diffuusiotensorikuvantaminen (DTI) mahdollistaa valkoisen aineen arvioinnin, koska se kykenee kuvaamaan veden diffuusio-ominaisuuksia (Mori et ai., 1999). Normaalissa valkoisessa aineessa vesimolekyyleillä on suhteellinen liikkumisvapaus hermokuitujen kanssa. Niiden liikkeet ovat kuitenkin rajoitettuja eri alueilla, mikä johtaa valkoisen aineen diffuusioanisotropiaan. Vastaavasti diffuusioanisotropiaa on käytetty arvioitaessa kuidun muutoksen laajuutta valkoisessa aineessa (Chang et ai., 2010; Puig et ai., 2010). Useissa viimeaikaisissa tutkimuksissa on yritetty osoittaa DTI: n hyödyllisyys vaurioiden arvioinnissa potilailla, joilla on heikentynyt tietoisuus ja tiettyjen ARAS-ytimien liitettävyys ihmisen aivoissa (Voss et al., 2006; Perlbarg et ai., 2009; Tollard et ai., 2009; Tshibanda ym., 2009; Fernandez-Espejo ym., 2010, 2011; Newcombe ym., 2010; Edlow ym., 2012). ARAS: n koko rekonstruoinnista ja arvioinnista ihmisen aivoissa tiedetään kuitenkin vähän (Edlow et al., 2012).
Tässä tutkimuksessa yritimme rekonstruoida alempaa yksittäistä komponenttia DTI: n avulla. ARAS-arvo pontiinin radiotaajuudesta talamuksen intralaminaarisiin ytimiin normaalissa ihmisen aivoissa.
Materiaalit ja menetelmät
Kohteet
26 normaalia terveellistä koehenkilöä Tähän tutkimukseen rekrytoitiin (14 miestä, 12 naista; keski-ikä 31,85 ± 9,80 vuotta; vaihteluväli 20–50), joilla ei ollut aiemmin ollut neurologista sairautta. Kaikki koehenkilöt osallistuivat tähän tutkimukseen vapaaehtoisina ja antoivat kirjallisen suostumuksen ennen DTI-skannausta. Tutkimuksen hyväksyi sairaalamme institutionaalinen arviointilautakunta.
Diffuusiotensorikuva
DTI-tiedot saatiin käyttämällä 6-kanavaista pääkäämiä 1,5-T Philips Gyroscan Interassa ( Philips, Best, Alankomaat), jossa on yhden kuvan kaikusuunnallinen kuvantaminen. Kullekin 32 ei-kolineaarisesta diffuusioherkistävästä gradientista hankimme 67 vierekkäistä viipaletta yhdensuuntaisesti etu- ja takaosan komentojohdon kanssa. Kuvankäsittelyparametrit olivat seuraavat: hankintamatriisi = 96 × 96, rekonstruoitu matriisiksi = 128 × 128, näkökenttä = 221 mm × 221 mm, TR = 10 726 ms, TE = 76 ms, rinnakkaiskuvan vähennyskerroin (SENSE-kerroin) = 2, EPI-kerroin = 49 ja b = 1000 s / mm2, NEX = 1 ja viipaleen paksuus 2,3 mm (hankittu isotrooppisen vokselin koko 2,3 mm × 2,3 mm × 2,3 mm).
Todennäköinen kuitu Seuranta
Diffuusiopainotettujen kuvantamistietojen analyysi suoritettiin käyttämällä Oxford Center for Functional Magnetic Resonance Imaging of Brain (FMRIB) -ohjelmistokirjastoa (FSL; www.fmrib.ox.ac.uk/fsl). Affine-moniulotteista kaksiulotteista rekisteröintiä käytettiin pyörrevirrasta johtuvan pään liikeefektin ja kuvan vääristymien korjaamiseen. Kuidunseuranta suoritettiin todennäköisyystraktografisella menetelmällä, joka perustui monikuituiseen malliin, ja sitä sovellettiin nykyisessä tutkimuksessa hyödyntämällä FMRIB-diffuusiossa toteutettuja traktografiarutiineja (5000 virtaviivaista näytettä, 0,5 mm: n askelpituudet, kaarevuusrajat = 0,2) (Smith et al., 2004 ).Tässä tutkimuksessa käytetyn todennäköisyystraktografian etuihin kuuluu suurempi kestävyys melulle sekä kyky havaita reittejä terävämmillä kulmilla ja erottaa ristikkäiset kuidut (Behrens et ai., 2007; Winston et ai., 2011).
ARAS-reitti määritettiin valitsemalla kuidut, jotka kulkevat mielenkiinnon kohteena olevien siemenalueiden (ROI) ja kohde (lopetus) ROI: n läpi. Siementen ROI sijoitettiin poneiden RF: ään kolmoishermon sisääntulovyöhykkeen tasolla (Daube, 1986; Afifi ja Bergman, 2005). Mediaalisen lemniscuksen ja rubrospinaalisen alueen analyysi tehtiin radion radion vahvistamiseksi poneissa (kuvio 1A). Mediaalisen lemniscuksen analysoimiseksi siemenen ROI: t asetettiin anteromediaaliseen medulaan ja kohde-ROI sijoitettiin somatosensoriseen aivokuoreen (Hong et ai., 2010). Ruosteen selkärangan analysoimiseksi siementen ROI: t sijoitettiin punaiseen ytimeen ja kohde-ROI sijoitettiin medullan vastakkaiseen dorsolateraaliseen alueeseen (Monakowin alue) (Nathan ja Smith, 1982; Kwon et ai., 2011). Kohde-ROI annettiin talamuksen intralaminaarisissa ytimissä kommissuaalisen tason tasolla (Morel, 2007). Määritellessäsi talamuksen intralaminaariset ytimet, viittasimme aivojen atlasiin (Morel, 2007) (kuva 1A). 5000 siemenvoxelista muodostetusta näytteestä kontaktin tulokset visualisoitiin kynnysarvolla vähintään 1 virtaviivainen jokaisen vokselin läpi analysointia varten. Mitattiin ARAS-alemman yksittäisen komponentin murto-anisotropian (FA), keskimääräisen diffuusiokyvyn (MD) ja traktorin numero.
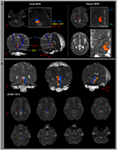
Kuva 1. (A) Kiinnostuksen kohteena olevat siemenalueet (ROI) on annettu pisteen verkkokalvomuodostuksessa (punainen väri). Kohde-ROI annetaan talamuksen intralaminaarisissa ytimissä kommissuaalisen tason tasolla. Talamuksen intralaminaaristen ytimien raja määriteltiin viittaamalla aivojen atlasen oppikirjaan (Morel, 2007). ML, mediaalinen lemniscus; RST, rintarauhasen alue; RF, retikulaarinen muodostuminen; AC, etukanava; PC, takaosa. (B) Rekonstruoidun nousevan retikulaarisen aktivointijärjestelmän polut näkyvät normaalin kohteen (26-vuotiaan miehen) jokaisella aivotasolla.
Tilastollinen analyysi
SPSS-ohjelmistoa (v.15.0; SPSS, Chicago, IL, USA) käytettiin tietojen analysointiin. Parillista t-testiä käytettiin ARAS: n DTI-parametrien arvojen erojen määrittämiseen oikean ja vasemman pallonpuoliskon välillä. Pearsonin korrelaatiotestiä käytettiin ARAS: n DTI-parametrien ja iän välisen korrelaation määrittämiseen. Tuloksia pidettiin merkittävinä, kun p-arvo oli < 0,05.
Tulokset
Rekonstruimme ARP: n alemman yksittäisen komponentin pontinin välillä Talamuksen RF- ja intralaminaariset ytimet. ARAS: n rekonstruoitu komponentti sai alkunsa pontin RF: stä, nousi mesencephalic tegmentumin läpi juuri takana punaisen ytimen läpi ja päättyi sitten talamuksen intralaminaarisiin ytimiin commissural-tason tasolla kaikissa kohteissa (kuva 1B). / p>
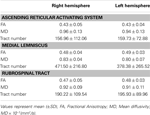
Taulukko 1. Nousevan retikulaarisen aktivointijärjestelmän diffuusiotensorikuvaparametrit .
Keskustelu
Nykyisessä tutkimuksessa rekonstruoimme yhden DTI: n avulla ARAS, ARAS: n alempi yksittäinen komponentti RF: stä talamukseen normaaleilla koehenkilöillä, vaikka ARAS koostuu ylimääräisistä aivorungon ytimistä, hypotalamuksesta, tyvi-aivoista ja talamokortikaalisista projektioista aivokuorelle. Valitsimme kaksi ROI: ta ARAS: n alemman yksittäisen komponentin rekonstruoimiseksi: siemen-ROI, joka oli ponssin RF RF kolmoishermon sisääntulovyöhykkeen tasolla (Daube, 1986; Afifi ja Bergman, 2005), ja kohde ROI, joka sisälsi talamuksen intralaminaariset ytimet (keskimmäiset lateraaliset ytimet, sentromedian / parafaskikulaariset ytimet ja paracentraaliset ytimet) kommissuaalisen tason tasolla (Morel, 2007). Aivorungon rostralinen osa kolmoishermon sisääntulovyöhykkeen yläpuolella tunnetaan nimellä ARAS; sen sijaan RF: n kaudaalinen osa on mukana sydämen ja hengityksen toimintaan liittyvässä motorisessa toiminnassa ja autonomisessa toiminnassa (Daube, 1986). Siksi sijoitimme siemenen ROI: n RF: ään kolmoishermon sisääntulovyöhykkeen tasolle. Sijoitimme kohde-ROI: n intralaminaarisiin ytimiin, jotka ovat ARAS: n päätuoleja epäspesifisten talamuksen ytimien joukossa. Siksi uskomme, että koska emme voineet sisällyttää muita ARAS: iin liittyviä talamuksen ytimiä, ARAS: n alempi yksittäinen komponentti, joka rekonstruoitiin tässä tutkimuksessa, ei ole ARAS: n koko alempi yksittäinen komponentti, vaan pääosa koko ARAS: n alempi yksittäinen komponentti.Tämän seurauksena ARAS: n alempi yksittäinen komponentti on peräisin pontin RF: stä, nousi punaisen ytimen takaosan mesencephalic tegmentumin läpi ja päättyi sitten talamuksen intralaminaarisiin ytimiin. Lisäksi ARAS: n rekonstruoidun alemman yksittäisen komponentin FA-, MD- ja traktaalinumeroiden arvot eivät eronneet merkittävästi oikean ja vasemman pallonpuoliskon välillä. Traktaaliluku määräytyy hermostoon sisältyvien vokselien lukumäärän perusteella (Kwak et ai., 2010). FA-arvo osoittaa valkean aineen mikrorakenteiden, kuten aksonit, myeliini ja mikrotubulit, suuntauksen ja eheyden, ja ADC-arvo osoittaa veden diffuusion suuruuden (Assaf ja Pasternak, 2008).
Useita tutkimuksia ovat osoittaneet DTI: n kliinisen hyödyllisyyden arvioimalla joitain ARAS: n alemman yksittäisen komponentin alueita RF: stä talamukseen tajunnan vajaatoimintaa sairastavilla potilailla (Perlbarg et al., 2009; Tollard et ai., 2009; Newcombe et ai., 2010; Fernandez-Espejo ym., 2011). Tollard et ai. (2009) raportoi DTI: n hyödyllisyydestä, joka tehtiin subakuutissa vaiheessa tuloksen ennustamiseksi 45 potilaalla, joilla oli vaikea TBI (traumaattinen aivovamma) (vastausten puuttuminen yksinkertaisista tilauksista). Tutkimuksessaan he mitasivat FA-arvon useilla supratentorialisilla ja infratentoriallisilla alueilla, mukaan lukien etuponit, posterioriset ponsit ja keskiaivot, ja osoittivat, että infratentorialisen ja supratentoriallisen FA: n väheneminen mahdollisti ennustaa epäedullisia tuloksia 1 vuosi TBI: stä. Perlbarg et ai. (2009), joka suoritti DTI-skannauksen 30 potilaalla ilman vastetta yksinkertaisille tilauksille vakavan TBI: n jälkeen, ilmoitti FA: n selvän vähenemisen mitattuna alemmassa pitkittäisessä fasciculuksessa, keskiaivoissa (aivopoika ja tegmentum), sisäisen kapselin takaosassa ja posterior corpus callosum epäedullisessa tulosryhmässä. Newcombe et ai. (2010) käytti DTI: tä valkeaainehäviön laajuuden ja sijainnin määrittelemiseen potilailla, jotka olivat vegetatiivisessa tilassa TBI: n seurauksena (seitsemän potilasta) ja potilailla, joilla oli iskeeminen-hypoksinen vamma (viisi potilasta). Poikkeavuuksia supratentorialisilla alueilla havaittiin molemmissa ryhmissä; sitä vastoin aivorungon poikkeavuuksia havaittiin vain TBI-ryhmässä. Fernandez-Espejo et ai. (2011) käytti DTI: tä 25 vegetatiivisen ja vähän tajuissa olevan potilaan neuropatologian erilaistamisessa. He päättelivät, että vähämerkityksiset potilaat ja vegetatiivisessa tilassa olevat potilaat poikkesivat toisistaan aivokuoren alla olevan valkoisen aineen ja talamuksen alueilla, mutta eivät näyttäneet eroavan aivorungosta. Äskettäisessä tutkimuksessa, jossa käytettiin korkean kulman tarkkuuden diffuusiokuvausta, Edlow et ai. (2012) raportoivat ARAS: n neuroanatomisesta liitettävyydestä ihmisen aivoissa sekä in vivo että ex vivo. He osoittivat, että tiettyjen ARAS-ytimien liitännät liittyivät kiihottumiseen ja talamisten ytimien yhteydet kiihottumisen modulointiin. RF ihmisen aivojen talamukseen. Uskomme, että käytetyt menetelmät ja tämän tutkimuksen tulokset voivat olla hyödyllisiä tutkijoille, jotka tutkivat ARAS: ta ihmisen aivoissa. Yksi tämän tutkimuksen rajoituksista on kuitenkin se, että emme pystyneet täysin selvittämään koko ARAS-järjestelmää, koska emme sisällyttäneet analyyseihimme muita talamuksen ja aivorungon ytimiä, jotka ovat myös mukana ARAS: ssa. Tarvitaan lisätutkimuksia havaintojemme kliinisestä hyödyllisyydestä sekä tutkimuksia ARAS: n ennusteista talamuksesta aivokuoreen.
Lausunto eturistiriidoista
Kirjoittajat julistavat että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Tätä työtä tuki DGIST R & Korean opetus-, tiede- ja teknologiaministeriön D-ohjelma (13-BD-0401).
Afifi, AK ja Bergman, RA (2005). Toiminnallinen neuroanatomia: Teksti ja Atlas. New York: Lange Medical Books / McGraw-Hill.
Morel, A. (2007). Ihmisen talamuksen ja basaaliganglian stereotaktinen atlas. New York: Informa Healthcare.