Półka
Funkcje peroksysomów
Peroksysomy zawierają co najmniej 50 różnych enzymów, które biorą udział w różnych szlakach biochemicznych w różnych typach komórek . Pierwotnie peroksysomy były definiowane jako organelle, które przeprowadzają reakcje utleniania prowadzące do produkcji nadtlenku wodoru. Ponieważ nadtlenek wodoru jest szkodliwy dla komórki, peroksysomy zawierają również enzym katalazę, który rozkłada nadtlenek wodoru, przekształcając go w wodę lub używając go do utleniania innego związku organicznego. W takich reakcjach utleniania w peroksysomach rozkładane są różne substraty, w tym kwas moczowy, aminokwasy i kwasy tłuszczowe. Szczególnie ważnym przykładem jest utlenianie kwasów tłuszczowych (ryc. 10.25), ponieważ stanowi główne źródło energii metabolicznej. W komórkach zwierzęcych kwasy tłuszczowe są utleniane zarówno w peroksysomach, jak i mitochondriach, ale w drożdżach i roślinach utlenianie kwasów tłuszczowych ogranicza się do peroksysomów.
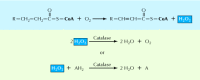
Rysunek 10.25
Utlenianie kwasów tłuszczowych w peroksysomach. Utlenianiu kwasu tłuszczowego towarzyszy produkcja nadtlenku wodoru (H2O2) z tlenu. Nadtlenek wodoru jest rozkładany przez katalazę, albo przez konwersję do wody, albo przez utlenianie innego organicznego (więcej …)
Oprócz zapewnienia przedziału dla reakcji utleniania, peroksysomy biorą udział w biosyntezie lipidów. W komórkach zwierzęcych cholesterol i dolichol są syntetyzowane w peroksysomach, a także w ER. W wątrobie peroksysomy biorą również udział w syntezie kwasów żółciowych, które pochodzą z cholesterolu. Ponadto peroksysomy zawierają enzymy wymagane do syntezy plazmalogenu – rodziny fosfolipidów, w której jeden z łańcuchów węglowodorowych jest połączony z glicerolem raczej wiązaniem eterowym niż estrowym (Rysunek 10.26). Plazmalogeny są ważnymi składnikami błon niektórych tkanek, szczególnie serca i mózgu, chociaż w innych są nieobecne.

Rysunek 10.26
Struktura plazmalogenu. Przedstawiony plazmalogen jest analogiczny do fosfatydylocholiny. Jednak jeden z łańcuchów kwasu tłuszczowego jest połączony z glicerolem wiązaniem eterowym, a nie estrowym.
Peroksysomy odgrywają w roślinach dwie szczególnie ważne role. Po pierwsze, peroksysomy w nasionach są odpowiedzialne za przemianę zmagazynowanych kwasów tłuszczowych w węglowodany, co ma kluczowe znaczenie dla dostarczania energii i surowców do wzrostu kiełkującej rośliny. Dzieje się to poprzez serię reakcji określanych jako cykl glioksylanowy, który jest wariantem cyklu kwasu cytrynowego (Rysunek 10.27). Peroksysomy, w których to ma miejsce, są czasami nazywane glioksysomami.
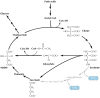
Rysunek 10.27
Cykl glioksalowy. Rośliny są zdolne do syntetyzowania węglowodanów z kwasów tłuszczowych poprzez cykl glioksylanowy, który jest wariantem cyklu kwasu cytrynowego (patrz Rysunek 2.34). Podobnie jak w cyklu kwasu cytrynowego, acetylo-CoA łączy się ze szczawiooctanem, tworząc (więcej …)
Po drugie, peroksysomy w liściach biorą udział w fotooddychaniu, które służy do metabolizowania produkt uboczny powstały podczas fotosyntezy (Rysunek 10.28). CO2 jest przekształcany w węglowodany podczas fotosyntezy poprzez szereg reakcji zwanych cyklem Calvina (patrz Rysunek 2.39). Pierwszym krokiem jest dodanie CO2 do pięciowęglowego cukru rybulozo-1,5-bisfosforanu, w wyniku czego powstają dwie cząsteczki 3-fosfoglicerynianu (po trzy węgle każda). Jednak zaangażowany enzym (karboksylaza rybulozobisfosforanu lub rubisco) czasami katalizuje dodanie O2 zamiast CO2, wytwarzając jedną cząsteczkę 3-fosfoglicerynianu i jedną cząsteczkę fosfoglikolanu (dwa atomy węgla). Jest to reakcja uboczna, a fosfoglikolan nie jest użytecznym metabolitem. Najpierw jest przekształcany w glikolan, a następnie przenoszony do peroksysomów, gdzie jest utleniany i przekształcany w glicynę. Glicyna jest następnie przenoszona do mitochondriów, gdzie dwie cząsteczki glicyny są przekształcane w jedną cząsteczkę seryny, z utratą CO2 i NH3. Seryna jest następnie zawracana do peroksysomów, gdzie jest przekształcana w glicerynian. Wreszcie glicerynian jest przenoszony z powrotem do chloroplastów, gdzie ponownie wchodzi w cykl Calvina. Wydaje się, że fotooddychanie nie jest korzystne dla rośliny, ponieważ zasadniczo jest odwrotnością fotosyntezy – zużywa się O2 i uwalnia się CO2 bez uzyskiwania ATP. Jednak okazjonalne wykorzystanie O2 zamiast CO2 wydaje się być nieodłączną właściwością rubisco, więc fotooddychanie jest ogólnym towarzyszeniem fotosyntezy. W ten sposób peroksysomy odgrywają ważną rolę, umożliwiając odzyskanie i wykorzystanie większości węgla w glikolanie.
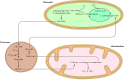
Rysunek 10.28
Rola peroksysomów w fotooddychaniu.Podczas fotosyntezy CO2 jest przekształcany w węglowodany w cyklu Calvina, który inicjowany jest przez dodanie CO2 do pięciowęglowego cukru rybulozo-1,5-bisfosforanu. Jednak czasami zaangażowany enzym (więcej …)