Frontiers in Human Neuroscience (Polski)
Wprowadzenie
Świadomość jest pobudzeniem i świadomością środowiska i siebie, co jest osiągane poprzez działanie wstępującego systemu aktywującego siatkowatego (ARAS) na pień mózgu i kora mózgowa (Daube, 1986; Paus, 2000; Zeman, 2001; Gosseries i in., 2011). ARAS składa się z kilku obwodów neuronalnych łączących pień mózgu z korą. Te połączenia neuronalne wywodzą się głównie z tworzenia siatkowatego (RF) pnia mózgu i przechodzą przez przekaźniki synaptyczne w jądrze międzywarstwowym wzgórza do kory mózgowej (Daube, 1986; Paus, 2000; Zeman, 2001; Afifi i Bergman, 2005; Gosseries i in., 2011). Ponadto, kilka jąder pnia mózgu (miejsce sinawe, szew grzbietowy, szew środkowy, jądro szypułkowo-czołowe, jądro okołobramkowe), nieswoiste jądra wzgórza, podwzgórze i podstawna część przodomózgowia są również objęte systemem ARAS (Aston-Jones i in., 2001 ; Parvizi i Damasio, 2003; Fuller i in., 2011). Dokładna ocena ARAS jest ważna dla diagnozy i leczenia pacjentów z zaburzeniami świadomości, takich jak pacjenci w stanie wegetatywnym lub z minimalną świadomością (Zeman, 2001; Gosseries i in., 2011).
W badaniach ARAS w ludzkim mózgu zastosowano konwencjonalne MRI mózgu, techniki funkcjonalnego neuroobrazowania, metody elektrofizjologiczne i spektroskopię MR (Parvizi i Damasio, 2003; Schiff, 2006; Tshibanda i in., 2009, 2010; Gawryluk i in. ., 2010). Ponieważ jednak ARAS nie można wyraźnie odróżnić od sąsiednich struktur neuronowych, dokładna identyfikacja i oszacowanie ARAS w ludzkim mózgu może być problematyczne przy stosowaniu tych metod. Natomiast obrazowanie tensorowe dyfuzji (DTI) pozwala na ocenę istoty białej ze względu na jej zdolność do obrazowania charakterystyk dyfuzji wody (Mori i in., 1999). W normalnej istocie białej cząsteczki wody mają względną swobodę ruchów równolegle do włókien nerwowych. Jednak ich ruchy są ograniczone w poprzek dróg, co prowadzi do anizotropii dyfuzyjnej istoty białej. W związku z tym do oceny stopnia zmiany włókien w istocie białej zastosowano anizotropię dyfuzyjną (Chang i in., 2010; Puig i in., 2010). W kilku ostatnich badaniach podjęto próbę wykazania przydatności DTI do oceny zmian chorobowych u pacjentów z upośledzoną świadomością i połączeniem określonych jąder ARAS w ludzkim mózgu (Voss i in., 2006; Perlbarg i in., 2009; Tollard i in., 2009; Tshibanda i in., 2009; Fernandez-Espejo i in., 2010, 2011; Newcombe i in., 2010; Edlow i in., 2012). Jednak niewiele wiadomo o całej rekonstrukcji i oszacowaniu ARAS w ludzkim mózgu (Edlow i in., 2012).
W obecnym badaniu, przy użyciu DTI, próbowaliśmy zrekonstruować dolną pojedynczą składową ARAS z mostu RF do jąder międzywarstwowych wzgórza w normalnym ludzkim mózgu.
Materiały i metody
Badani
Dwadzieścia sześć zdrowych osób (14 mężczyzn, 12 kobiet; średni wiek 31,85 ± 9,80 lat; zakres 20–50) bez historii choroby neurologicznej. Wszyscy badani uczestniczyli w tym badaniu jako ochotnicy i przedstawili pisemną zgodę przed poddaniem się skanowaniu DTI. Badanie zostało zatwierdzone przez komisję rewizyjną naszego szpitala.
Obraz Tensora Dyfuzji
Dane DTI uzyskano przy użyciu 6-kanałowej cewki głowowej na 1,5-tonowym aparacie Philips Gyroscan Intera ( Philips, Best, Holandia) z jednostrzałowym obrazowaniem echo-planarnym. Dla każdego z 32 niekoliniowych gradientów uczulających na dyfuzję uzyskano 67 przylegających skrawków równoległych do przedniej linii spoidła tylnego. Parametry obrazowania były następujące: matryca akwizycji = 96 × 96, odtworzona do matrycy = 128 × 128, pole widzenia = 221 mm × 221 mm, TR = 10726 ms, TE = 76 ms, współczynnik redukcji obrazowania równoległego (współczynnik SENSE) = 2, współczynnik EPI = 49 i b = 1000 s / mm2, NEX = 1 i grubość warstwy 2,3 mm (uzyskany izotropowy rozmiar woksela 2,3 mm × 2,3 mm × 2,3 mm).
Włókno probabilistyczne Śledzenie
Analiza danych obrazowania ważonego dyfuzyjnie została przeprowadzona przy użyciu biblioteki oprogramowania Oxford Center for Functional Magnetic Resonance Imaging of the Brain (FMRIB) (FSL; www.fmrib.ox.ac.uk/fsl). Do korekcji efektu ruchu głowy i zniekształcenia obrazu wywołanego prądami wirowymi zastosowano afiniczną wieloskalową rejestrację dwuwymiarową. Śledzenie włókien zostało przeprowadzone przy użyciu probabilistycznej metody traktografii opartej na modelu wielowłóknowym i zastosowane w obecnym badaniu z wykorzystaniem procedur traktograficznych zaimplementowanych w FMRIB Diffusion (5000 próbek opływowych, długość kroku 0,5 mm, progi krzywizny = 0,2) (Smith i in., 2004 ).Zalety traktografii probabilistycznej, która została wykorzystana w tym badaniu, to większa odporność na szum, a także zdolność wykrywania ścieżek o ostrzejszych kątach i rozróżniania krzyżujących się włókien (Behrens i in., 2007; Winston i in., 2011).
Ścieżka ARAS została określona przez wybór włókien przechodzących przez obszary zainteresowania nasion (ROI) i docelowe (końcowe) ROI. Nasiono ROI umieszczono na RF mostu na poziomie strefy wejścia nerwu trójdzielnego (Daube, 1986; Afifi i Bergman, 2005). Przeprowadzono analizę przyśrodkowego lemniscus i rubrospinal, aby potwierdzić granicę RF na moście (ryc. 1A). W celu analizy przyśrodkowej lemniscus, obszary ROI nasion umieszczono na rdzeniu przednio-przyśrodkowym, a docelowy obszar ROI umieszczono na korze somatosensorycznej (Hong et al., 2010). W celu analizy przewodu rubrospinalowego, obszary ROI nasion umieszczono na jądrze czerwonym, a docelowy obszar ROI umieszczono na przeciwległym grzbietowo-bocznym regionie rdzenia (obszar Monakowa) (Nathan i Smith, 1982; Kwon i wsp., 2011). Docelowy zwrot z inwestycji został podany na jądrach wewnątrzwarstwowych wzgórza na poziomie płaszczyzny spoidłowej (Morel, 2007). Definiując jądra wewnątrzwarstwowe wzgórza, odnieśliśmy się do atlasu mózgu (Morel, 2007) (ryc. 1A). Spośród 5000 próbek wygenerowanych z woksela nasion, wyniki kontaktu były wizualizowane przy minimalnym progu 1 przepuszczanym przez każdy woksel do analizy. Zmierzono wartości ułamkowej anizotropii (FA), średniej dyfuzyjności (MD) i liczby dróg dolnego pojedynczego składnika ARAS.
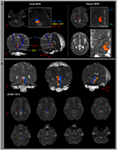
Rysunek 1. (A) Badane obszary nasienne (ROI) podano na formacji siatkowatej mostu (kolor czerwony). Docelowy obszar ROI jest podawany na jądrach międzywarstwowych wzgórza na poziomie płaszczyzny spoidłowej. Granicę międzywarstwowych jąder wzgórza określono na podstawie podręcznika atlasu mózgu (Morel, 2007). ML, przyśrodkowy lemniscus; RST, przewód rubrospinal; RF, formacja siatkowata; AC, spoidło przednie; PC, spoidło tylne. (B) Ścieżki zrekonstruowanego wstępującego systemu aktywacji siatkowatej są pokazane na każdym poziomie mózgu u normalnego podmiotu (26-letni mężczyzna).
Analiza statystyczna
Do analizy danych użyto oprogramowania SPSS (wersja 15.0; SPSS, Chicago, IL, USA). Do określenia różnicy wartości parametrów DTI ARAS pomiędzy prawą a lewą półkulą zastosowano sparowany test t-Studenta. Do określenia korelacji między parametrami DTI ARAS a wiekiem zastosowano test korelacji Pearsona. Wyniki uznawano za istotne, gdy wartość p wynosiła < 0,05.
Wyniki
Zrekonstruowaliśmy dolną pojedynczą składową ARAS między mostem RF i jądra wewnątrzwarstwowe wzgórza. Zrekonstruowany komponent ARAS pochodził z mostu RF, wznosił się przez śródmózgowiowe nakrywki tuż za jądrem czerwonym, a następnie kończył się w jądrach wewnątrzwarstwowych wzgórza na poziomie płaszczyzny spoidłowej u wszystkich badanych (ryc. 1B).
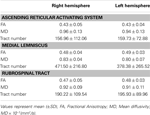
Tabela 1. Parametry obrazowania za pomocą tensora dyfuzji wstępującego systemu aktywującego siatkowatego .
Dyskusja
W obecnym badaniu, używając DTI, zrekonstruowaliśmy jedną z głównych ścieżek ARAS, dolny pojedynczy składnik ARAS od RF do wzgórza u zdrowych osób, chociaż ARAS składa się z dodatkowych jąder pnia mózgu, podwzgórza, podstawnego przodomózgowia i wypustek wzgórzowo-korowych do kory mózgowej. Wybraliśmy dwa obszary ROI do rekonstrukcji dolnego pojedynczego komponentu ARAS: ROI nasion, który był RF mostu na poziomie strefy wejścia nerwu trójdzielnego (Daube, 1986; Afifi i Bergman, 2005) oraz docelowy ROI, który obejmował jądra wewnątrzwarstwowe wzgórza (jądra centralne boczne, jądra centromedian / parafascicular i jądra paracentral) na poziomie płaszczyzny spoidłowej (Morel, 2007). Przednia część RF pnia mózgu powyżej strefy wejścia nerwu trójdzielnego jest znana jako ARAS; przeciwnie, ogonowa część RF jest zaangażowana w funkcje motoryczne i autonomiczne związane z czynnością serca i układu oddechowego (Daube, 1986). Dlatego umieściliśmy ROI nasion w RF na poziomie strefy wejścia nerwu trójdzielnego. Docelowy obszar zainteresowania ROI umieściliśmy w jądrach międzywarstwowych, które są głównymi jądrami ARAS wśród niespecyficznych jąder wzgórza. Dlatego uważamy, że ponieważ nie mogliśmy uwzględnić innych jąder wzgórzowych związanych z ARAS, dolny pojedynczy składnik ARAS, który został zrekonstruowany w obecnym badaniu, nie jest całym dolnym pojedynczym składnikiem ARAS, ale główną częścią cały dolny pojedynczy składnik ARAS.W konsekwencji, dolny pojedynczy składnik ARAS wywodził się z mostu RF, wznosił się przez śródmózgowiową powłokę z tyłu jądra czerwonego, a następnie kończył się w jądrach międzywarstwowych wzgórza. Ponadto wartości FA, MD i liczby dróg zrekonstruowanej dolnej pojedynczej składowej ARAS nie różniły się istotnie między prawą a lewą półkulą. Liczba dróg zależy od liczby wokseli znajdujących się w przewodzie nerwowym (Kwak i in., 2010). Wartość FA wskazuje stopień kierunkowości i integralności mikrostruktur istoty białej, takich jak aksony, mielina i mikrotubule, a wartość ADC wskazuje wielkość dyfuzji wody (Assaf i Pasternak, 2008).
Kilka badań wykazali kliniczną przydatność DTI, szacując niektóre obszary dolnej pojedynczej składowej ARAS od RF do wzgórza u pacjentów z zaburzeniami świadomości (Perlbarg i in., 2009; Tollard i in., 2009; Newcombe i in., 2010; Fernandez-Espejo i in., 2011). Tollard i in. (2009) opisali przydatność DTI, którą wykonano na podostrym etapie w przewidywaniu wyniku u 45 pacjentów z ciężkim TBI (urazowym uszkodzeniem mózgu) (brak odpowiedzi na proste polecenia). W swoich badaniach zmierzyli wartość FA w kilku obszarach nadnamiotowych i podnamiotowych, w tym w przednim, tylnym moście i śródmózgowiu, i wykazali, że zmniejszenie przednamiotowego i nadnamiotowego FA, z wyjątkiem tylnego mostu, pozwoliło przewidzieć niekorzystne wyniki 1 rok od TBI. Perlbarg i in. (2009), którzy przeprowadzili badanie DTI u 30 pacjentów z brakiem odpowiedzi na proste polecenia po ciężkim TBI, donieśli o zdecydowanym zmniejszeniu FA mierzonego w pęczku podłużnym dolnym, śródmózgowiu (nasadka mózgowa i nakrywka), kończynie tylnej torebki wewnętrznej i tylne ciało modzelowate w grupie z niekorzystnym wynikiem. Newcombe i in. (2010) zastosowali DTI do scharakteryzowania zakresu i lokalizacji utraty istoty białej u pacjentów, którzy byli w stanie wegetatywnym wtórnym do TBI (7 pacjentów) i pacjentów z urazem niedokrwienno-hipoksycznym (5 pacjentów). Nieprawidłowości w obszarach nadnamiotowych zaobserwowano w obu grupach; z kolei nieprawidłowości pnia mózgu obserwowano tylko w grupie TBI. Fernandez-Espejo i in. (2011) zastosowali DTI w różnicowaniu neuropatologii 25 wegetatywnych i mało świadomych pacjentów. Doszli do wniosku, że pacjenci z minimalną świadomością i pacjenci w stanie wegetatywnym różniły się podkorową substancją białą i regionami wzgórza, ale nie wydawało się, aby różniły się pniem mózgu. W niedawnym badaniu wykorzystującym obrazowanie dyfuzyjne o wysokiej rozdzielczości kątowej Edlow i wsp. (2012) opisali łączność neuroanatomiczną ARAS w ludzkim mózgu, zarówno in vivo, jak i ex vivo. Wykazali, że połączenia określonych jąder ARAS były zaangażowane w pobudzenie, a połączenia jąder wzgórza – z modulacją pobudzenia.
Podsumowując, używając DTI, zrekonstruowaliśmy dolny pojedynczy składnik ARAS z RF do wzgórza w ludzkim mózgu. Uważamy, że zastosowana metodologia i wyniki tego badania mogą być pomocne dla naukowców badających ARAS w ludzkim mózgu. Jednak jednym z ograniczeń tego badania jest to, że nie byliśmy w stanie w pełni wyjaśnić całego systemu ARAS, ponieważ nie uwzględniliśmy w naszej analizie innych jąder wzgórza i pnia mózgu, które są również zaangażowane w ARAS. Potrzebne są dalsze badania przydatności klinicznej naszych ustaleń, a także badania nad projekcją ARAS od wzgórza do kory mózgowej.
Oświadczenie o konflikcie interesów
Autorzy deklarują że badanie zostało przeprowadzone przy braku jakichkolwiek powiązań handlowych lub finansowych, które można by zinterpretować jako potencjalny konflikt interesów.
Podziękowania
Praca była wspierana przez DGIST R & D Program Ministerstwa Edukacji, Nauki i Technologii Korei (13-BD-0401).
Afifi, AK i Bergman, RA (2005). Neuroanatomia funkcjonalna: tekst i atlas. Nowy Jork: Lange Medical Books / McGraw-Hill.
Morel, A. (2007). Atlas stereotaktyczny ludzkiego wzgórza i zwojów podstawy. Nowy Jork: Informa Healthcare.